
Blog|Articles|July 19, 2019
Prion-like Molecular Mechanisms in Alzheimer Disease
Author(s)Lary C. Walker, PhD
Although Alzheimer disease is not infectious by any common definition of the term, research over the past 20 years has confirmed long-standing speculation that the molecular mechanism driving neurodegeneration is fundamentally the same in Alzheimer disease and the prototypical infectious proteopathy-prion disease.
Advertisement
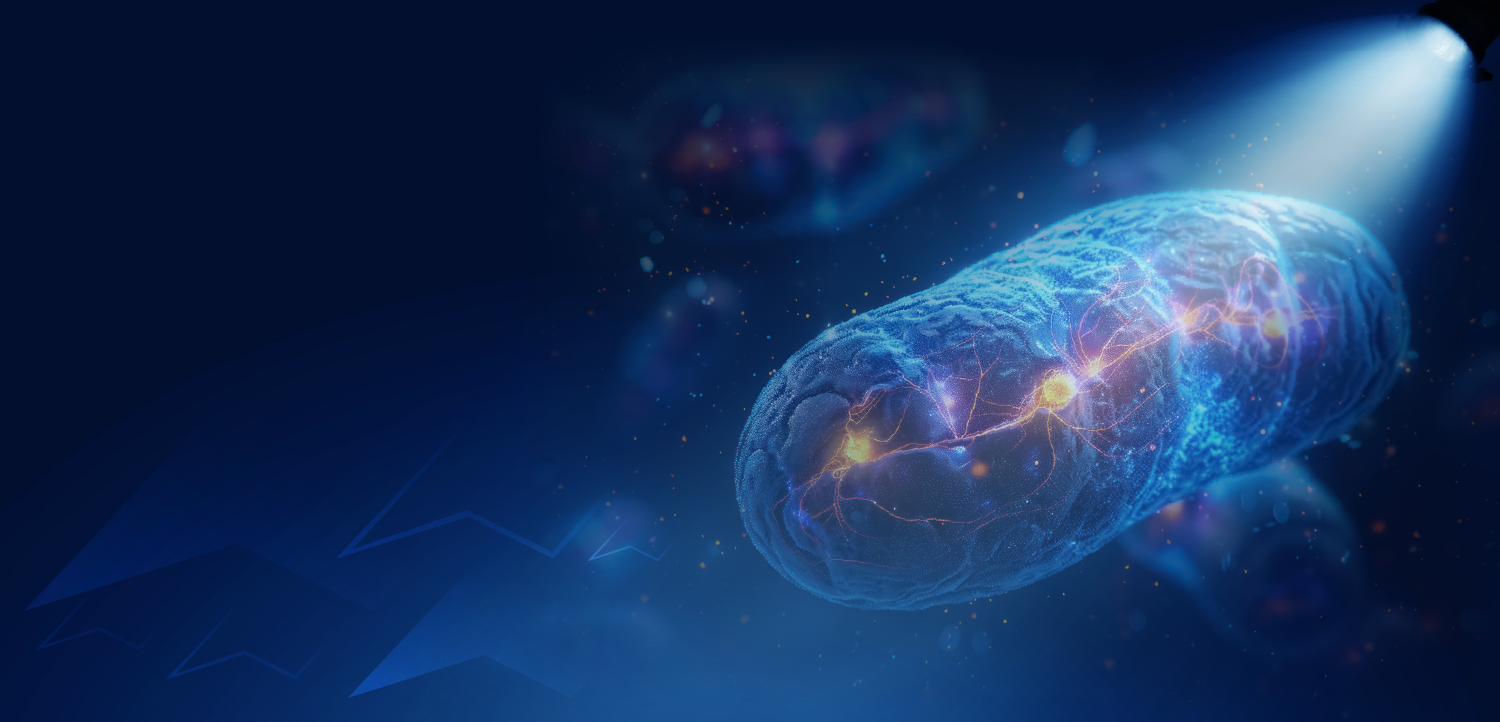
Although Alzheimer disease is not infectious by any common definition of the term, research over the past 20 years has confirmed long-standing speculation that the molecular mechanism driving neurodegeneration is fundamentally the same in Alzheimer disease and the prototypical infectious proteopathy-prion disease. This mechanism is seeded protein aggregation, essentially a crystallization-like process by which certain proteins misfold into a highly stable conformation that compels like proteins to misfold and stick to one another (FIGURE 1).
The transmissibility of prions results from the transfer of proteinaceous infectious particles-prions-from one organism to another.1 Although considerable attention has been given to this unorthodox type of infectivity, most human prion diseases, similar to Alzheimer disease,
The prion paradigm of disease
The prion diseases are extraordinary in that they can be infectious, genetic, or idiopathic in origin. They are
Most human prion diseases arise endogenously, presumably with the spontaneous misfolding and aggregation of the prion protein. Years or even decades can pass between the initiation of the disease process and the emergence of signs and symptoms. As the aberrant prion proteins accumulate in the nervous system, they form deposits that are visible under the light-microscope (FIGURE 2), along with small, stealthy assemblies called oligomers that are thought to be especially toxic to cells. Relatively late in the pathogenic process, the deterioration and demise of neurons causes brain function to decline precipitously, with death often occurring within a year of clinical onset. At autopsy, the prion-laden brain is riddled with vacuoles, giving affected regions a spongiform appearance.
Alzheimer disease
Alzheimer disease bears intriguing similarities to prion diseases; it involves a chronic, degenerative and ultimately fatal disease mechanism in which misfolded proteins accumulate in the brain.2 The earliest event is the structural corruption of the Aβ protein that forms senile plaques and cerebral amyloid angiopathy, followed by the misfolding and hyperphosphorylation of Tau protein that forms neurofibrillary tangles (FIGURE 3). The proliferation of these two corrupted proteins is considered the defining feature of Alzheimer disease, and like the prion protein aberran Aβ and Tau also give rise to cytotoxic oligomers.4,5
Another similarity is that, in both disorders, the pathogenic cascade begins in the brain many years before the first clear behavioral impairments emerge.4 Alzheimer can be genetic or idiopathic in origin, but it has not been shown to be infectious. Nevertheless, a compelling case can be made that the proteins that define Alzheimer as a distinct disease entity-Aβ and Tau-misfold, aggregate, and spread through the brain in a manner similar to that of prions.
The prion-like properties of misfolded Aβ and Tau
In 1998, my colleague Mathias Jucker and I began a
We then undertook a series of experiments demonstrating that, point by point, the characteristics of Aβ seeds are identical to those of prions (TABLE). Specifically, these studies showed that Aβ seeds:
1. Are protein-only agents that induce disease in susceptible hosts
2. Spread within and to the brain
3. Exist in multiple sizes and shapes (called strains)
4. Can resist inactivation by heat and formaldehyde
Importantly, a number of research groups now have shown that Tau seeds also share these molecular features with prions (Table).7-10
Accidental Aβ seeding in humans
From the late 1950s until the mid-1980s, certain young people with short stature were treated with intramuscular injections of growth hormone derived from pituitary glands collected from human cadavers at autopsy.11,12 Large batches of glands were homogenized to extract the hormone, which nicely accelerated the children’s growth. Many years later, however, a subset of the treated patients developed Creutzfeldt-Jakob disease, probably because some pituitaries in the homogenized lots came from patients who had died with prion disease. (In 1985, a safer, recombinant form of growth hormone was introduced.)
Hypothesizing that the large batches also were likely to include pituitaries from persons with Alzheimer disease, in 2015
Fully developed Alzheimer pathology-including marked AB-proteopathy and tauopathy-has not been established in any of these instances. Whether recipients of cadaveric growth hormone or dura mater transplants are at a higher risk of Alzheimer disease in the future is not known. However, the transmission of cerebral amyloid angiopathy has important clinical ramifications because vascular amyloid increases the risk for cerebral hemorrhage.17,18
Implications of the expanded prion paradigm
Experimental and iatrogenic evidence for the seedability of AB aggregation is a reminder that surgical instruments-particularly those used in neurosurgery-should be pristine. While there is no indication that Alzheimer disease can be conveyed by blood transfusion, Creutzfeldt-Jakob has, in very rare instances, been so transmitted.19 Because advancing age is a strong risk factor for Alzheimer disease and other proteopathies, it thus may not be unreasonable to consider age limits on the donation of blood and other biologics. In addition, the role of other possible environmental and physiological risk factors needs clarification, as these might be viable therapeutic targets.20
Perhaps the most important implication of the expanded prion paradigm, however, is that this fundamental molecular mechanism-seeded protein aggregation-in all likelihood drives the pathogenesis of a remarkable array of complex diseases, ranging from Alzheimer disease and Creutzfeldt-Jakob disease to frontotemporal dementia, amyotrophic lateral sclerosis, Parkinson disease, Lewy body dementia, Huntington disease, non-cerebral amyloidoses, and quite possibly many others.21 As a unifying molecular principle, the prion paradigm can serve as the polestar for the development of therapeutic strategies for many diseases that continue to defy effective treatment.
Dr Walker is Marie and E. R. Snelling Professor, Department of Neurology and Research Professor, Yerkes National Primate Research Center,
References:
1. Prusiner SB.
2. Walker LC. Prion-like mechanisms in Alzheimer disease. Handbk Clin Neurol. 2018;153:303-319.
3. Will RG, Ironside JW.
4. Jack CR, Jr, Bennett DA, Blennow K, et al. NIA-AA Research Framework: toward a biological definition of Alzheimer disease. Alzh Dement. 2018;14:535-562.
5. Jucker M, Walker LC. Propagation and spread of pathogenic protein assemblies in neurodegenerative diseases. Nat Neurosci. 2018;21:1341-1349.
6. Jucker M, Walker LC.
7. Clavaguera F, Tolnay M, Goedert M. The prion-like behavior of assembled Tau in transgenic Mice. Cold Spring Harb Perspect Med. 2017;7(10).
8. Gibbons GS, Lee VMY, Trojanowski JQ. Mechanisms of cell-to-cell transmission of pathological Tau: a review. JAMA Neurol. 2019;76:101-108.
9. Holmes BB, Diamond MI. Cellular models for the study of prions. Cold Spring Harb Perspect Med. 2017;7(2).
10. Aoyagi A, Condello C, Stöhr J, et al. AB and tau prion-like activities decline with longevity in the Alzheimer's disease human brain. Sci Transl Med. 2019;11(490).
11. Blizzard RM. History of growth hormone therapy. Indian J Pediatr. 2012;79:87-91.
12. Rudge P, Jaunmuktane Z, Adlard P, et al. Iatrogenic CJD due to pituitary-derived growth hormone with genetically determined incubation times of up to 40 years. Brain. 2015;138(Pt 11):3386-3399.
13. Jaunmuktane Z, Mead S, Ellis M, et al.
14. Ritchie DL, Adlard P, Peden AH, et al. Amyloid-beta accumulation in the CNS in human growth hormone recipients in the UK. Acta Neuropathol. 2017;134:221-240.
15. Frontzek K, Lutz MI, Aguzzi A, et al. Amyloid-beta pathology and cerebral amyloid angiopathy are frequent in iatrogenic Creutzfeldt-Jakob disease after dural grafting. Swiss Med Wkly. 2016;146:w14287.
16. Hamaguchi T, Taniguchi Y, Sakai K, et al. Significant association of cadaveric dura mater grafting with subpial Ab deposition and meningeal amyloid angiopathy. Acta Neuropathol. 2016;132:313-315.
17. Purro SA, Farrow MA, Linehan J, et al. Transmission of amyloid-beta protein pathology from cadaveric pituitary growth hormone. Nature. 2018;564:415-419.
18. Auriel E, Greenberg SM. The pathophysiology and clinical presentation of cerebral amyloid angiopathy. Curr Atheroscler Rep. 2012;14:343-350.
19. Brown P, Brandel JP, Sato T, et al. Iatrogenic Creutzfeldt-Jakob disease, final assessment. Emerg Infect Dis. 2012;18:901-907.
20. Walker LC, Lynn DG, Chernoff YO. A standard model of Alzheimer disease? Prion. 2018;12:261-265.
21. Prusiner SB, Ed. Prion Diseases. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2017.
22. Mirbaha H, Chen D, Morazova OA, et al. Inert and seed-competent tau monomers suggest structural origins of aggregation. Elife. 2018;7.
23. Duyckaerts C, Sazdovitch V, Ando K, et al. Neuropathology of iatrogenic Creutzfeldt-Jakob disease and immunoassay of French cadaver-sourced growth hormone batches suggest possible transmission of tauopathy and long incubation periods for the transmission of Abeta pathology. Acta Neuropathol. 2018;135:201-212.
Advertisement
Related Content
Advertisement




Latest CME










Advertisement
Advertisement
Trending on NeurologyLive - Clinical Neurology News and Neurology Expert Insights
1
Managing Infection Risk and Safety Monitoring with Anti-CD20 MS Disease Modifying Therapy
2
Long-Term Efficacy of Anti-CD20 Therapies in MS: What the Evidence Shows
3
Former NFL Running Back Chris Johnson Diagnosed With ALS at 40
4
Where Tolebrutinib Fits: Weighing the EU Approval, FDA Divergence, and the Road Ahead in nrSPMS
5