
|Articles|June 16, 2021
- June 2021
- Volume 4
- Issue 3
Myostatin Inhibition in Amyotrophic Lateral Sclerosis
Author(s)Jennifer S. Sun, PhD
ALS has only recently been associated with causative or disease-modifying mutations in 20 genes that encode proteins with diverse functions, with promising therapeutic targets including proteins in pathways that regulate protein homeostasis.
Advertisement
AMYOTROPHIC LATERAL SCLEROSIS (ALS) is a fatal neurodegenerative disorder characterized by progressive death of motor neurons, which causes muscle atrophy that results in disability.1,2 Although no cure exists for ALS, available therapeutics have focused on managing symptoms to prolong survival and improve quality of life,2 while experimental treatments for ALS seek to stop or reverse motor neuron damage and muscle atrophy to slow or stop disease progression.
The etiology of ALS is largely unknown; symptoms vary by individual and are dependent on the associated genes and presence of comorbidities.1 Moreover, these symptoms can initially be subtle and mimic those of other neurodegenerative disorders, making early diagnosis challenging.2 Approximately 90% of ALS cases are of sporadic origin, whereas 10% are familial.2,3 This multifactorial disease has only recently been associated with causative or disease-modifying mutations in 20 genes that encode proteins with diverse functions, including RNA metabolism, vesicle trafficking, proteasomal function, and oxidative stress).4 Promising ALS therapeutic targets, therefore, include proteins in pathways that regulate protein homeostasis.5
Myokines are one such family of cytokines that can function as autocrine, paracrine, or endocrine stimuli.4,6 Muscle contraction induces the biosynthesis and release of myokines, whereas reduced physical activity impairs the myokine response.4,6 Understanding the biological implications of myokines can help inform development of therapeutics to halt or reverse the physiological effects of progressive brain disorders.
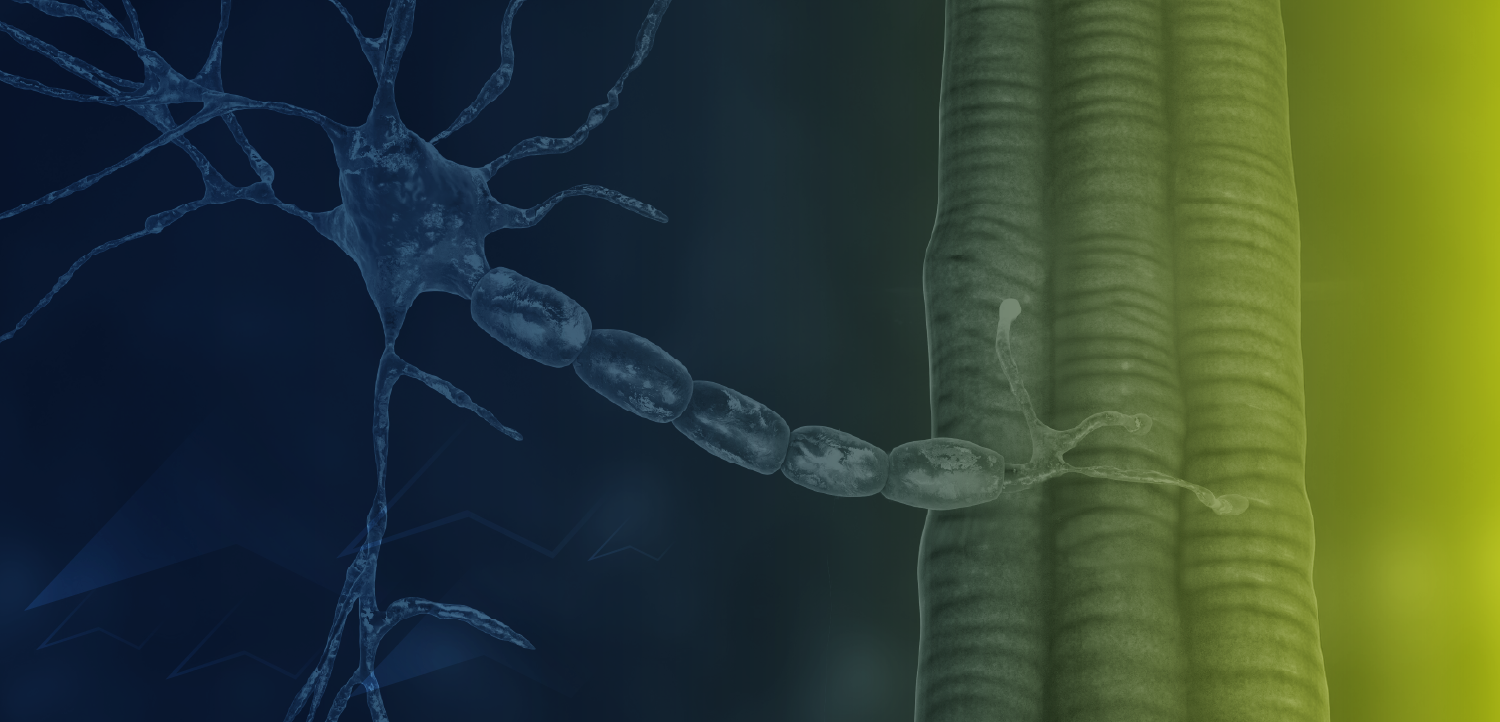
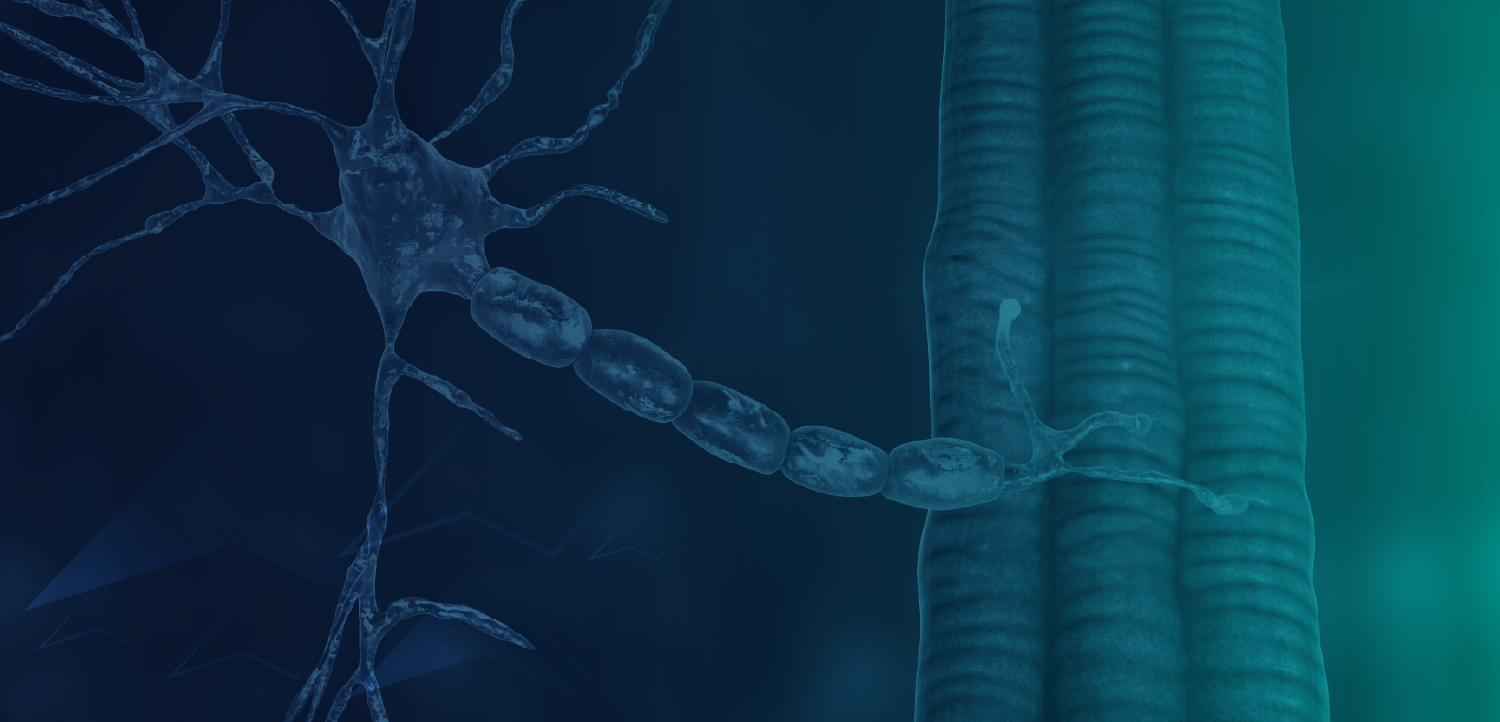
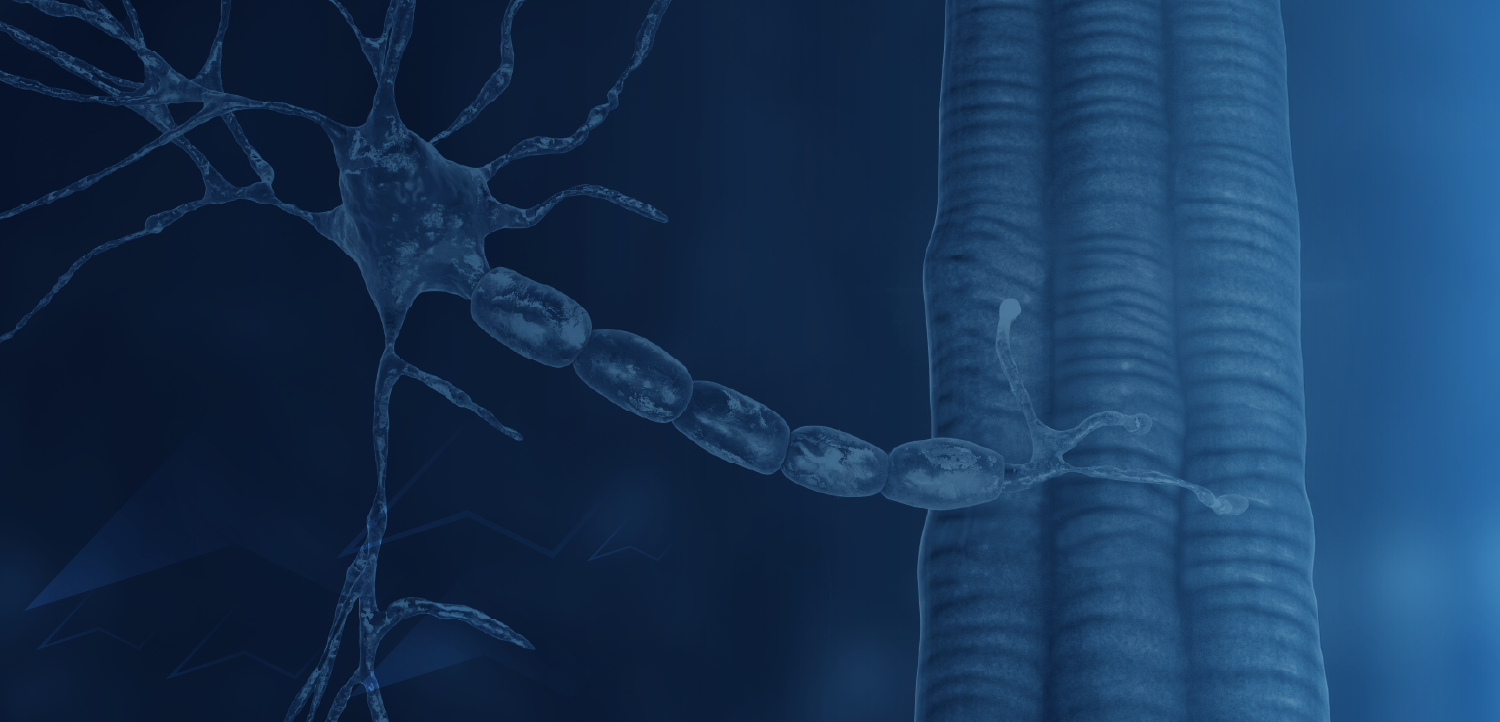
Myostatin, a myokine in the TGF-β superfamily of ligands (also known as growth/differentiation factor 8; GDF8), disrupts protein synthesis and negatively regulates skeletal muscle mass.7-9 Characteristic of the structure of TGF-β family members, immature myostatin exhibits an N-terminal prodomain with a C-terminal signaling domain.9,10 A proteolytic cleavage removes the prodomain, but it remains noncovalently associated with the signaling domain, maintaining it in an inhibited latent state.7,9,10 The mature signaling domain becomes activated when the prodomain is fully removed via a second cleavage, which can be caused by fluctuations in pH.9 Activated myostatin can then bind its cell surface receptor complex consisting of a type I receptor (ALK4/5) and a type II receptor (ACTRIIA/B), thereby acting as a potent negative regulator of muscle mass.6-8
Genetic or chemical disruption of myostatin has been shown to substantially promote skeletal muscle growth.6,10 A high abundance of myostatin, compared with levels of its natural inhibitor follistatin, is positively correlated with muscle wasting and fat loss in mouse models.6,7,10 Conversely, myostatin inhibition consistently increases muscle mass in animal models of a variety of muscle disorders.6-8 Myostatin downregulation is also achievable naturally through sustained regular exercise of moderate intensity.11
Myostatin inhibition is under investigation as a means to reverse progressive muscle wasting (FIGURE).8,10 Most investigational antimyostatin drug candidates are antibodies designed to block receptor signaling. However, receptor recognition surfaces of TGF-β family members share a high degree of similarity.10 Toxicity could result from antibody promiscuity.10,11 Moreover, these protein targets are known to bind to a number of ligands, so therapies may not specifically block myostatin action.10
To overcome this high ligand homology, prodomain-targeting therapeutics (neutralizing antibodies, short interfering RNA, small molecules, etc) are currently under development. Myostatin-specific drugs promise increased specificity and tolerability.11 These novel therapeutic approaches aim to block myostatin maturation from the latent state.10 For example, exogenous follistatin (FS-344) delivered by a single adeno‐associated virus (AAV1) injection has long-lasting benefits in dystrophic muscles: reduced inflammation accompanied by increased muscle size and strength.11 GASP1, which shares binding domain sequence similarity with follistatin, is capable of complexing with latent and active myostatin.10,11 Investigators have recently identified the amino acid residues in the prodomain that maintain myostatin in the latent state.9 This knowledge may help inform design of effective prodomain-targeting molecules. Gene therapy may provide lasting results and overcome potential immune responses toward recombinant protein injection.11
Antisense oligonucleotide (AON) therapy has been validated in mouse models, where a resulting decrease in activated myostatin expression corresponded with a significant increase in muscle mass.11,12 Combinatorial therapy with AON and AAV-based therapies has been associated with adverse effects (AEs), including thrombocytopenia.11 However, AON therapy has some advantages over traditional AAV-based gene therapies: (1) the possibility of local or systemic administration, (2) easily tailored dosing regimen and downregulation intensity, (3) low toxicity and immunogenicity, and (4) no risk of uncontrolled insertion into the genome.11,12
Myostatin inhibition appears to be a useful therapeutic approach for attenuating and reversing the muscle atrophy characteristic of patients with ALS.10 However, studies have shown that oxidoreductases such as superoxide dismutase (SOD1) accelerate disease progression despite increased muscle mass or strength.13 Edaravone (Radicava; Mitsubishi Tanabe Pharma America) is an FDA-approved ALS therapy that counteracts excessive oxidative stress, although it is not specific and has been linked to serious AEs in nontarget organs.3 Other investigational therapeutics for ALS include low-dose administration of an iron chelator such as Deferiprone to offset oxidative stress in motor pathways.14
There is still much to learn about the etiology of ALS, both familial and sporadic. Until then, symptomatic treatment remains focused on helping preserve quality of life and prolong survival.
For correspondence: [email protected]
New York Genome Center, New York, NY
REFERENCES
1. Scaricamazza S, Salvatori I, Giacovazzo G, et al. Skeletal-muscle metabolic reprogramming in ALS-SOD1G93A mice predates disease onset and is a promising Therapeutic Target. iScience. 2020;23(5):101087. doi:10.1016/j.isci.2020.101087
2. Štětkářová I, Ehler E. Diagnostics of amyotrophic lateral sclerosis: up to date. Diagnostics (Basel). 2021;11(2). doi:10.3390/diagnostics11020231
3. Cruz MP. Edaravone (Radicava): a novel neuroprotective agent for the treatment of amyotrophic lateral sclerosis. P T. 2018;43(1):25-28.
4. Halon-Golabek M, Borkowska A, Herman-Antosiewicz A, Antosiewicz J. Iron metabolism of the skeletal muscle and neurodegeneration. Front Neurosci. 2019;13:165. doi:10.3389/fnins.2019.00165
5. Malik R, Wiedau M. Therapeutic approaches targeting protein aggregation in amyotrophic lateral sclerosis. Front Mol Neurosci. 2020;13:98. 10.3389/fnmol.2020.00098
6. Lee JH, Jun HS. Role of myokines in regulating skeletal muscle mass and function. Front Physiol. 2019;10:42. doi:10.3389/fphys.2019.00042
7. Sumner CJ, Wee CD, Warsing LC, et al. Inhibition of myostatin does not ameliorate disease features of severe spinal muscular atrophy mice. Hum Mol Genet. 2009;18(17):3145-3152. doi:10.1093/hmg/ddp253
8. Smith RC, Lin BK. Myostatin inhibitors as therapies for muscle wasting associated with cancer and other disorders. Curr Opin Support Palliat Care. 2013;7(4):352-360. doi:10.1097/SPC.0000000000000013
9. Walker RG, McCoy JC, Czepnik M, et al. Molecular characterization of latent GDF8 reveals mechanisms of activation. Proc Natl Acad Sci. 2018;115(5):E866-E875. doi:10.1073/pnas.1714622115
10. Pirruccello-Straub M, Jackson J, Wawersik S, et al. Blocking extracellular activation of myostatin as a strategy for treating muscle wasting. Sci Rep. 2018;8(1):2292. doi:10.1038/s41598-018-20524-9
11. Mariot V, Joubert R, Hourdé C, et al. Downregulation of myostatin pathway in neuromuscular diseases may explain challenges of anti-myostatin therapeutic approaches. Nat Commun. 2017;8(1):1859. doi:10.1038/s41467-017-01486-4
12. Liu CM, Yang Z, Liu CW, et al. Myostatin antisense RNA-mediated muscle growth in normal and cancer cachexia mice. Gene Ther. 2008;15(3):155-160. doi:10.1038/sj.gt.3303016
13. Prudencio M, Hart PJ, Borchelt DR, Andersen PM. Variation in aggregation propensities among ALS-associated variants of SOD1: Correlation to human disease. Hum Mol Genet. 2009;18(17):3217-3226. doi:10.1093/hmg/ddp260
14. Moreau C, Danel V, Devedjian JC, et al. Could conservative iron chelation lead to neuroprotection in amyotrophic lateral sclerosis? Antioxid Redox Signal. 2018;29(8):742-748. doi:10.1089/ars.2017.7493
Articles in this issue
about 5 years ago
The State of Coma Care: A Call for Actionabout 5 years ago
Love and a Lab Coat: One Couple’s Quest to Treat Alzheimer Diseaseabout 5 years ago
Biohaven’s Zavegepant Aims to Expand CGRP Migraine Treatment Optionsabout 5 years ago
Refining the Approach to Seizure Rescue Medicationsabout 5 years ago
Do We Finally Have a Useful “Sed Rate” for Multiple Sclerosis?about 5 years ago
Holding On to Hope in Huntington Diseaseabout 5 years ago
Aducanumab Approved for Alzheimer Disease TreatmentAdvertisement
Related Content
Advertisement




Latest CME









Advertisement
Advertisement
Trending on NeurologyLive - Clinical Neurology News and Neurology Expert Insights
1
Rethinking Clinical Trial Design and the Immune System in Multiple Sclerosis: Ahmed Obeidat, MD, PhD
2
AAIC 2026 Preview: David Wolk, MD, on Biomarkers, Disease Heterogeneity, and What's Next in Alzheimer Research
3
Looking Beyond Gene Therapy for Treating Charcot-Marie-Tooth Disease: Carla Taveggia, PhD
4
Rituximab Noninferior to Ocrelizumab in Newly Diagnosed Relapsing MS, Head-to-Head Trial Finds
5