
|Articles|June 14, 2022
- June 2022
- Volume 5
- Issue 3
Endoplasmic Reticulum and/or Mitochondrial-Dependent Neuronal Degeneration in ALS
Author(s)Jennifer S. Sun, PhD
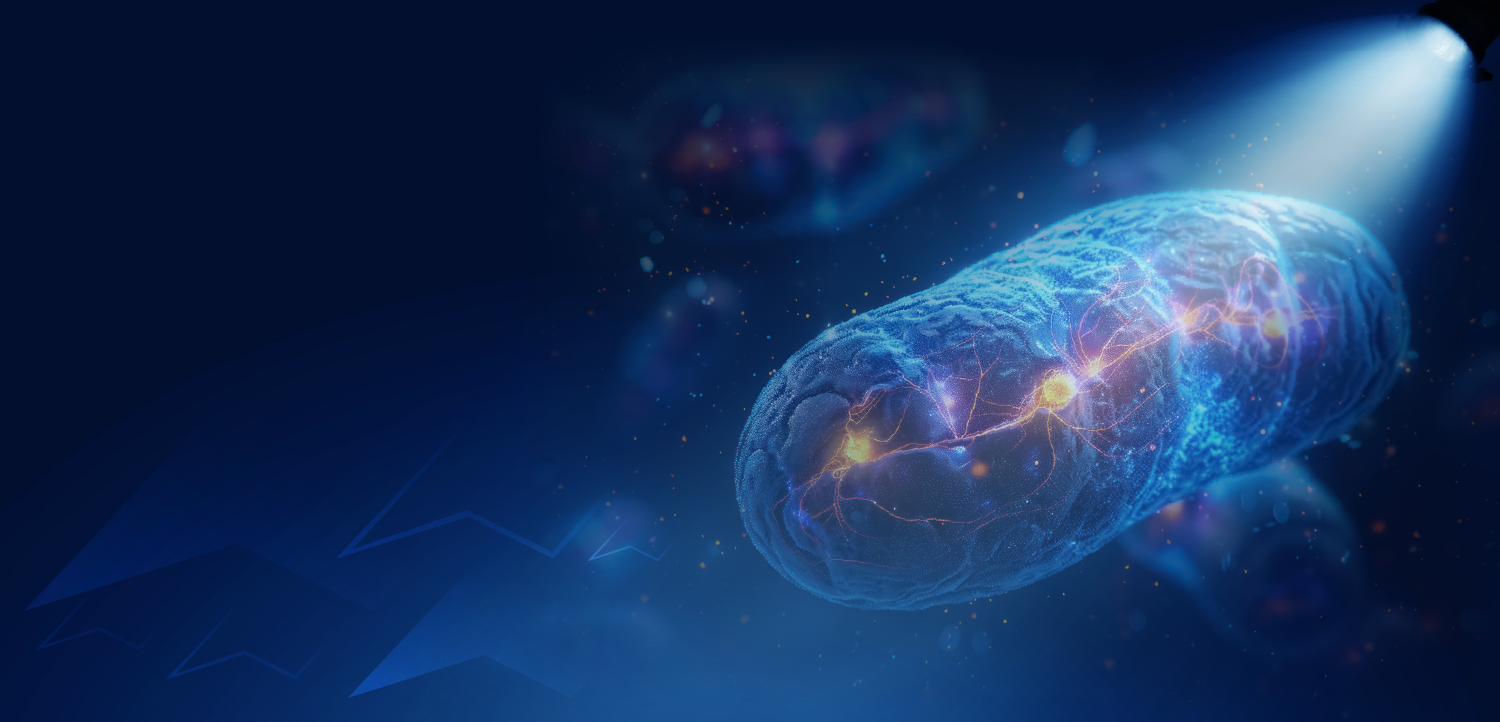
Given the importance of mitochondrial and endoplasmic reticulum function in ALS, the disruption of intracellular mitochondria-endoplasmic reticulum contacts presents yet another avenue for neuronal degeneration—perhaps the primary point of underlying dysfunction.
Advertisement
AMYOTROPHIC LATERAL SCLEROSIS (ALS) is an aggressive neurodegenerative disease with an average age of onset between 40 and 60 years.1,2 ALS presents with progressive degeneration of upper and lower motor neurons, which eventually culminates in lethal respiratory failure within 2 to 5 years of diagnosis.1,2 ALS can result from genetic and nongenetic factors, although there is currently no clear biological link between ALS-associated gene mutations and neural defects.1,3 In fact, only 10% of cases are familial; the remaining 90% are sporadic with unknown etiology.1,2 The leading hypothesis is that aberrant protein expression and aggregation impacts mitochondrial function, which in turn reduces calcium metabolism and triggers reactive oxygen species (ROS) overproduction that underlies motor neuron demise.1,3,4 Dysregulation of other proteostasis-associated pathways—axonal transport, autophagy, nucleocytoplasmic transport, endoplasmic reticulum (ER) stress—also contributes to ALS initiation and progression as part of a vicious feed-forward cycle.1,4
The maintenance of ionic gradients, particularly for intracellular calcium signaling, is essential to fulfill the energetic requirements of motor neurons.5 The ER is the primary site of protein and lipid biosynthesis and intracellular calcium storage, whereby the ER controls calcium concentration, which powers oxidative phosphorylation in the mitochondria.5 Calcium uptake by mitochondria is, in turn, dependent on the mitochondrial membrane potential, which is initially generated by calcium flux and is maintained by the electron transport chain in respiration.1,3,4 Communication between the ER and the mitochondria is essential to coordinate protein and lipid biosynthesis, calcium storage, and adenosine triphosphate (ATP) synthesis.1,5 Aberrant formation of phospholipids by the ER, and protein aggregate–triggered pore formation, can alter ER and/or mitochondrial function.1,3,5 For example, TAR DNA-binding protein 43 (TDP43) and fused in sarcoma (FUS) aggregates alter the mitochondrial proteome and impair buffering against oxidative stress in the cytoplasm.1 Moreover, required membrane interactions may become lost because of protein aggregation.1
In fact, the suspected underlying mechanism of familial ALS appears to mirror that of central nervous system disorders: accumulation of unfolded protein aggregates, which leads to oxidative stress that compromises organelle function and results in motor neuron degeneration.1,3,6 Of the 20 genes that have been associated with ALS to date, the major gene attributed to familial ALS, unsurprisingly, encodes superoxide dismutase 1 (SOD1), which normally scavenges mitochondrial ROS to reduce cellular toxicity; however, accumulated mutations in SOD1 result in gain of function effects which cause SOD1 aggregates to form within mitochondria of motor neurons to create oxidative stress.1 Mutant SOD1 and TDP43 genes have also been identified in patients with sporadic ALS and have been connected to the observed dysregulation of oxidative phosphorylation, overall mitochondrial respiration, and ATP production.1,4 Another prominent mutation associated with familial ALS is a noncoding GGGGCC repeat in the chromosome 9 open reading frame 72 (C9ORF72) gene, which encodes a chaperone in the electron transport chain, suggesting a role of RNA processing and nucleocytoplasmic transport in evoking toxicity.1,4 Overall, a disrupted redox environment increases the risk of 1) insufficient mitochondrial ATP production, which is an essential energetic requirement for motor neurons; and 2) shutdown of ER-driven protein synthesis, which impairs the production of phospholipids required for neuronal function.1,2,4,5
Given the importance of mitochondrial and ER function in ALS, disruption of intracellular mitochondria-ER contacts (MERCs, FIGURE) presents yet another avenue for neuronal degeneration, which could perhaps be the primary point of dysfunction underlying ALS.1,5,7 MERCs are generated, in part, by a tether formed between the outer membrane vesicle-associated membrane protein–associated protein B (VAPB) and protein tyrosine phosphatase–interacting protein 51 (PTPIP51), and maintained by regulatory proteins including the ubiquitin-specific chaperone valosin-containing protein (VCP).1,3-5 Importantly, synaptic activity is directly correlated with the presence of MERCs; increased synaptic activity increases MERC formation, whereas the loss of MERCs reduces synaptic transmission.4 Loss of MERCs is therefore detrimental to calcium communication and ATP production and can ultimately lead to neuronal death.4 It is estimated that 15% to 20% of the mitochondrial surface is connected to the ER.5 Studying the ER and mitochondria as a functional unit could thus reveal more about the underlying genetic disease mechanism in ALS.4 Indeed, calcium miscommunication between the ER and mitochondria has recently emerged as a major factor in the loss of calcium homeostasis in ALS.4
The regions of close physical contact between the 2 organelles (mitochondria-associated membranes; MAMs) facilitate molecular signaling and physical exchange that underlie processes such as calcium signaling, phospholipid metabolism, and autophagy.1,3,7 Altogether, MAMs control mitochondrial metabolism, buffer oxidative stress, and contribute to regulation of inflammation and cell death.1,3,7 A primary function of VAPB-PTPIP51 tethers is to facilitate inositol triphosphate receptor (IP3R)–mediated delivery of calcium from ER stores to mitochondria.7,8 The MAM-associated chaperone protein sigma 1 receptor (Sigma1R) is also involved in lipid export and calcium signaling through IP3R regulation.3 Accordingly, Sigma1R levels are significantly lower in patients with ALS than in healthy populations.3 Loss of VAPB-PTPIP51 has been correlated with reduction of synaptic transmission as a result of impaired calcium flux and reduced ATP production, which in turn cause myopathy.1,2,5
The formation of ROS, potentially due to malfunctioning SOD1, can also damage these MAMs by forming pores that then alter MAM function and interaction.1 In mutant SOD1 (mutSOD1) mice, mutSOD1 aggregates have been shown to localize to the MAM and inhibit MERCs.9 When Sigma1R was also ablated, further impairment of mitochondrial function was recorded, suggesting a functional interplay among SOD1, Sigma1R, and MAMs in ALS.9 It is, therefore, no surprise that ER ultrastructure is altered and MERCs are lost in mouse ALS populations.3 C9ORF72 is also enriched in MERCs, whereby mutant C9ORF72 containing expanded repeats can cause damage prior to disease onset by specifically disrupting VAPB-PTPIP51 tethers and impairing IP3R-mediated calcium delivery,2,8 although no overt differences in these organelles were observed in C9ORF72-mutant mice.8 The causative relationship between protein aggregates and ALS remains to be comprehensively investigated.1
Currently, no cure exists for ALS, partially because of its complex etiology and the inability to prioritize cell functions for drug discovery.7,8,9 Understanding the time line of disease development, particularly the events that lead up to ER-mitochondria dysfunction, can further the development of antioxidant strategies that can promote healthy ER-mitochondria interactions to possibly prevent ALS altogether.3,7,8 Effective targets for ALS therapeutics could include components of MAMs, particularly for the purpose of correcting defects in calcium homeostasis, which is a prominent feature of ALS.5 For instance, compensating for mutations that disrupt ER-mitochondrial signaling (such as VAPB) has been effective in repairing VAPB-PTPIP51 tethering and ER-mitochondria signaling in rodent models and promises to be a broadly therapeutic strategy for neurodegenerative diseases.7,8 Agonists of Sigma1R have also been successful in protecting motor neurons and muscle activity in presymptomatic ALS mutSOD1 mice and in mice with spontaneous neurodegeneration, suggesting that Sigma1R agonists can improve calcium homeostasis regardless of comorbidities.5,7 ER stress modulators such as Salubrinal have also been shown to restore calcium homeostasis in SOD1-mutant mice with defective Sigma1R.5 Activating or blocking IP3R could also modulate calcium homeostasis.5 Further downstream, correcting mitochondrial NAD+ levels could improve neuronal health.6 However, something to consider is that dysregulation of ER-mitochondrial signaling could be a secondary effect to other damaged features, in which case it is important to define the primary driver of ALS.7 Moreover, studies in human subjects, rather than rodent ALS models, will reveal whether MERCs are indeed perturbed in ALS and whether MERC damage can be used not only as an early diagnostic feature, but as a potential drug target.7
For correspondence: [email protected]
Rutgers University, New Brunswick, NJ
REFERENCES
1. Chen J, Bassot A, Giuliani F, Simmen T. Amyotrophic Lateral Sclerosis (ALS): Stressed by dysfunctional mitochondria-endoplasmic reticulum contacts (MERCs). Cells. 2021;10(7):1789. doi:10.3390/cells10071789
2. Singh T, Jiao Y, Ferrando LM, et al. Neuronal mitochondrial dysfunction in sporadic amyotrophic lateral sclerosis is developmentally regulated. Sci Rep. 2021;11(1):18916. doi:10.1038/ s41598-021-97928-7
3. Manfredi G, Kawamata H. Mitochondria and endoplasmic reticulum crosstalk in amyotrophic lateral sclerosis. Neurobiol Dis. 2016;90:35-42. doi:10.1016/j.nbd.2015.08.004
4. Dafinca R, Barbagallo P, Talbot K. The Role of mitochondrial dysfunction and ER stress in TDP-43 and C9ORF72 ALS. Front Cell Neurosci. 2021;15. Accessed April 22, 2022. https://www.frontiersin. org/article/10.3389/fncel.2021.653688
5. Bernard-Marissal N, Chrast R, Schneider BL. Endoplasmic reticulum and mitochondria in diseases of motor and sensory neurons: a broken relationship? Cell Death Dis. 2018;9(3):1-16. doi:10.1038/ s41419-017-0125-1
6. Gautam M, Gunay A, Chandel NS, Ozdinler PH. Mitochondrial dysregulation occurs early in ALS motor cortex with TDP-43 pathology and suggests maintaining NAD+ balance as a therapeutic strategy. Sci Rep. 2022;12(1):4287. doi:10.1038/s41598-022-08068-5
7. Lau DHW, Hartopp N, Welsh NJ, et al. Disruption of ER−mitochondria signalling in fronto-temporal dementia and related amyotrophic lateral sclerosis. Cell Death Dis. 2018;9(3):1-8. doi:10.1038/ s41419-017-0022-7
8. Gomez-Suaga P, Mórotz GM, Markovinovic A, et al. Disruption of ER-mitochondria tethering and signalling in C9orf72-associated amyotrophic lateral sclerosis and frontotemporal dementia. Aging Cell. 2022;21(2):e13549. doi:10.1111/acel.13549
9. Candelise N, Salvatori I, Scaricamazza S, et al. Mechanistic insights of mitochondrial dysfunction in amyotrophic lateral sclerosis: an update on a lasting relationship. Metabolites. 2022;12(3):233. doi:10.3390/metabo12030233
Articles in this issue
about 4 years ago
Updates in Dravet Syndrome: Entering a New Eraabout 4 years ago
Achieving Greater Diversity in Dementia Research: A Call to Actionabout 4 years ago
New Treatments in Migraine: An In-Depth Reviewabout 4 years ago
FDA Approves Oral Version of Edaravone for ALSAdvertisement
Related Content
Advertisement




Latest CME










Advertisement
Advertisement
Trending on NeurologyLive - Clinical Neurology News and Neurology Expert Insights
1
Roche Unveils PrevenTRON, a Phase 3 Prevention Trial of Trontinemab in Cognitively Unimpaired Individuals at High Risk of Alzheimer Decline
2
Alzheimer's Association Launches PROTECT-Cog Combination Prevention Trial
3
Sanofi's Oral TREM2 Agonist SAR448851 Advances to Phase 2 TREMHANCE Trial in Early Alzheimer Disease
4
FDA Accepts BLA for Z-Rostudirsen in DMD, Sets January PDUFA Date
5